L'AFC félicite ses 3 lauréats du prix de thèse 2018. Les lauréats ont présenté leurs travaux lors du colloque de l'Association Française de Cristallographie du 10 au 13 juillet 2018 à Lyon.
Le prix AFC - Mention Biologie a été attribué à Clément Charenton, Laboratoire de Biochimie « Molecular Bases and Regulation of Protein Biosynthesis » à l'Ecole Polytechnique à Palaiseau, thèse soutenue le 20 septembre 2016 sous la direction de Marc Graille. Titre - Études structurales et fonctionnelles de la dégradation de la coiffe des ARNm.
Le prix AFC - Mention Chimie a été attribué à Cassandre Kouvatas, Institut des Sciences Chimiques de Rennes, thèse soutenue le 28 septembre 2017 sous la direction de Thierry Bataille, Eric Le Fur et Laurent Le Pollès. Titre - Caractérisation structurale multiéchelle operando de catalyseurs industriels de type phosphate de vanadium.
Le prix AFC - Mention Physique a été attribué à Virginie Théry, Centre Européen de la Céramique (IRCer) à Limoges, thèse soutenue le 10 novembre 2017 sous la direction d'Alexandre Boulle, Aurelian Crunteanu et Jean-Christophe Orlianges. Titre - Étude de la microstructure et des transitions de phases électroniques et cristallines de couches épitaxiales de VO2 déposées sur différents substrats.
Le prix Biologie (Clément Charenton):
 J'ai effectué ma thèse au sein du Laboratoire de Biochimie de l'École Polytechnique sous la supervision du Dr Marc Graille et en collaboration avec l'équipe du Dr Bertrand Séraphin (IGBMC, Illkirch). Mes travaux se sont focalisés sur les mécanismes moléculaires de la dégradation de la coiffe protectrice des ARN messagers (ARNm) eucaryotes.
J'ai effectué ma thèse au sein du Laboratoire de Biochimie de l'École Polytechnique sous la supervision du Dr Marc Graille et en collaboration avec l'équipe du Dr Bertrand Séraphin (IGBMC, Illkirch). Mes travaux se sont focalisés sur les mécanismes moléculaires de la dégradation de la coiffe protectrice des ARN messagers (ARNm) eucaryotes. La dégradation des ARNm occupe une position centrale dans le contrôle de l'expression génétique en permettant aux cellules d'adapter leur répertoire de transcrits à leur environnement ou à divers évènements cellulaires (Parker 2012). L'élimination de la coiffe protégeant l'extrémité 5' des ARNm, appelée « decapping », constitue une étape cruciale de la dégradation des ARNm eucaryotes. Elle est communément considérée comme l'ultime point de contrôle avant la destruction totale du message (Hsu and Stevens 1993). Ainsi, des dérégulations dans le processus de decapping peuvent avoir des effets délétères sur le développement normal des cellules, se traduisant chez l'humain par des retards mentaux (Ahmed et al. 2015).
La réaction de decapping est catalysée par l'enzyme Dcp2 qui génère une molécule de m7GDP et un ARN présentant une extrémité 5' phosphate libre. Cette dernière peut ensuite être reconnue comme substrat par l'exonucléase cytoplasmique Xrn1 (Jinek, Coyle, and Doudna 2011) qui dégrade alors rapidement l'ensemble de l'ARNm. L'activité de Dcp2 est extrêmement contrôlée par de nombreux facteurs protéiques organisés au sein d'un réseau d'interaction complexe (Arribas-Layton et al. 2013). Depuis plus de vingt ans, de nombreuses études ont permis d'identifier les différents acteurs de la machinerie de decapping. Cependant, les bases moléculaires de la régulation de l'activité de Dcp2 par ses régulateurs protéiques restaient extrêmement mal comprises. Pendant ma thèse, grâce à une combinaison d'approches structurales (cristallographie aux rayons X) et fonctionnelles (biochimie, biophysiques, biologie moléculaire), j'ai pu décrire précisément les principaux mécanismes de contrôle de l'activité de decapping.
Pendant mes travaux, je me suis tout d'abord attaché à caractériser le domaine C-terminal du facteur de decapping Pat1 dont la structure avait été résolue précédemment par mon équipe d'accueil (Fourati et al. 2014).
La dégradation des ARNm est typiquement initiée par le raccourcissement de la queue poly(A) lors d'un processus appelé déadénylation. Les ARNm sont alors dits " oligo-adénylés ". Ils sont la cible du complexe Pat1-Lsm1-7 qui interagit à leur extrémité 3' (He and Parker 2001) (Fig A.) tout en, et de manière contre-intuitive, favorisant le decapping à leur extrémité 5' (Bouveret 2000). J'ai élucidé les bases moléculaires de cette fonction en démontrant que Pat1 recrute directement le complexe de decapping via plusieurs motifs linéaires appelés HLMs présents au sein de son extension C-terminale déstructurée (Fig B.). J'ai caractérisé précisément ces interactions par fluorescence et j'ai résolu les structures cristallographiques du domaine C-terminal de Pat1 en complexe avec trois de ces HLMs. J'ai finalement mis en évidence que Pat1 interagit aussi avec un HLM présent dans l'extension C-terminale de l'exonucléase Xrn1 (Fig E.). En résumé, cette étude a révélé le rôle de Pat1 dans le recrutement successif des machineries de decapping et de dégradation 5'-3' aux ARNm oligo-adenylés (Charenton et al. 2017).
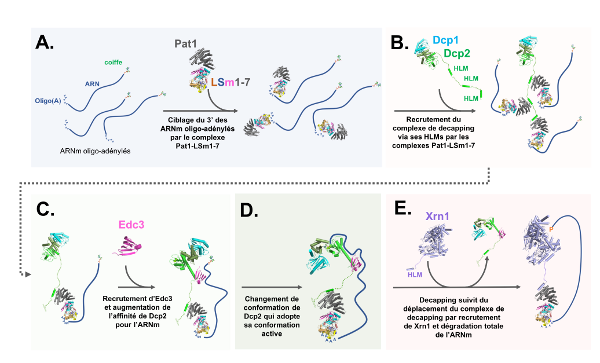
J'ai ensuite déterminé les structures cristallographiques du complexe de decapping Dcp1-Dcp2 seul ou en présence de son activateur Edc3 et d'un analogue de coiffe (m7GDP) (Charenton et al. 2016). La comparaison de ces deux structures a révélé que le facteur Edc3 stimule le decapping en induisant un changement de conformation de Dcp2 qui a pour effet d'augmenter son affinité pour les ARNm (Fig C.). De plus, la structure du complexe Dcp1-Dcp2-Edc3-m7GDP a dévoilé, pour la première fois, les bases structurales de la reconnaissance et du clivage de la coiffe par Dcp2. Finalement, cette étude a permis de comprendre comment le complexe de decapping adopte une conformation active lors de la catalyse, expliquant ainsi toutes les données fonctionnelles accumulées sur le sujet depuis près de dix ans (Coller 2016) (Fig D.).
En résumé, mes travaux ont permis d'établir un modèle pour le recrutement du complexe de decapping aux ARNm oligo-adenylés, pour son activation catalytique et pour la coordination de son activité avec celle de l'exonucléase Xrn1 qui élimine les ARNm une fois privés de leur coiffe (Fig A. à E.).
Références
Ahmed, Iltaf, Rebecca Buchert, Mi Zhou, Xinfu Jiao, Kirti Mittal, Taimoor I. Sheikh, Ute Scheller, et al. 2015. "Mutations in DCPS and EDC3 in Autosomal Recessive Intellectual Disability Indicate a Crucial Role for MRNA Decapping in Neurodevelopment." Human Molecular Genetics 24 (11): 3172-80. https://doi.org/10.1093/hmg/ddv069.
Arribas-Layton, Marcos, Donghui Wu, Jens Lykke-Andersen, and Haiwei Song. 2013. "Structural and Functional Control of the Eukaryotic MRNA Decapping Machinery." Biochimica Et Biophysica Acta 1829 (6-7): 580-89. https://doi.org/10.1016/j.bbagrm.2012.12.006.
Bouveret, E. 2000. "A Sm-like Protein Complex That Participates in MRNA Degradation." The EMBO Journal 19 (7): 1661-71. https://doi.org/10.1093/emboj/19.7.1661.
Charenton, Clément, Claudine Gaudon-Plesse, Zaineb Fourati, Valerio Taverniti, Régis Back, Olga Kolesnikova, Bertrand Séraphin, and Marc Graille. 2017. "A Unique Surface on Pat1 C-Terminal Domain Directly Interacts with Dcp2 Decapping Enzyme and Xrn1 5'-3' MRNA Exonuclease in Yeast." Proceedings of the National Academy of Sciences, October, 201711680. https://doi.org/10.1073/pnas.1711680114.
Charenton, Clément, Valerio Taverniti, Claudine Gaudon-Plesse, Régis Back, Bertrand Séraphin, and Marc Graille. 2016. "Structure of the Active Form of Dcp1-Dcp2 Decapping Enzyme Bound to m(7)GDP and Its Edc3 Activator." Nature Structural & Molecular Biology 23 (11): 982-86. https://doi.org/10.1038/nsmb.3300.
Coller, Jeff. 2016. "MRNA Decapping in 3D." Nature Structural & Molecular Biology 23 (11): 954-56. https://doi.org/10.1038/nsmb.3315.
Fourati, Zaineb, Olga Kolesnikova, Régis Back, Jenny Keller, Clément Charenton, Valerio Taverniti, Claudine Gaudon Plesse, et al. 2014. "The C-Terminal Domain from S. Cerevisiae Pat1 Displays Two Conserved Regions Involved in Decapping Factor Recruitment." PloS One 9 (5): e96828. https://doi.org/10.1371/journal.pone.0096828.
He, W., and R. Parker. 2001. "The Yeast Cytoplasmic LsmI/Pat1p Complex Protects MRNA 3' Termini from Partial Degradation." Genetics 158 (4): 1445-55.
Hsu, C L, and A Stevens. 1993. "Yeast Cells Lacking 5'-->3' Exoribonuclease 1 Contain MRNA Species That Are Poly(A) Deficient and Partially Lack the 5' Cap Structure." Molecular and Cellular Biology 13 (8): 4826-35.
Jinek, Martin, Scott M. Coyle, and Jennifer A. Doudna. 2011. "Coupled 5? Nucleotide Recognition and Processivity in Xrn1-Mediated MRNA Decay." Molecular Cell 41 (5): 600-608. https://doi.org/10.1016/j.molcel.2011.02.004.
Parker, Roy. 2012. "RNA Degradation in Saccharomyces Cerevisae." Genetics 191 (3): 671-702. https://doi.org/10.1534/genetics.111.137265.
Le prix Chimie (Cassandre Kouvatas):

Mon travail a d'abord consisté en une optimisation soigneuse des synthèses afin d'isoler sélectivement différents polymorphes. De plus, ces solides, pulvérulents par nature, présentent une cristallinité variable et un certain degré de désordre local, statique et/ou dynamique. Leur caractérisation impose donc une approche multiéchelle. Outre les techniques d'analyses thermiques et de microscopies associées, nous avons utilisé une combinaison de méthodes de caractérisation : des études en diffraction des rayons X (étude à l'échelle globale, quantification), en RMN du solide (sensibilité aux faibles variations de structure, extraction d'informations locales de géométrie, de désordre ou de dynamique) et des calculs théoriques basés sur des hypothèses structurales (confirmation ou non, ou éléments de réponse dans les descriptions structurales développées). L'intérêt de nos travaux réside non seulement dans leur caractère ex situ (température ambiante), mais aussi in situ (en température et sous atmosphère choisie). Plus particulièrement, des études operando (proches des conditions catalytiques) nous ont permis de caractériser les VPO au plus près des conditions réactives du processus industriel.
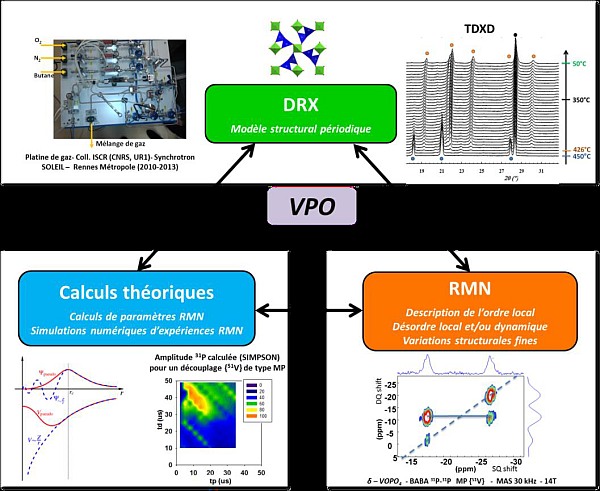
Une part importante du travail de thèse a porté sur des développements méthodologiques en RMN du solide. En effet, d'une structure à l'autre, les déplacements chimiques isotropes 31P des VOPO observés sont très proches, avec des largeurs à mi-hauteurs relativement importantes [5,6], du fait d'interactions scalaires hétéronucléaires entre un spin ½ (ici 31P) et un noyau quadrupolaire de spin demi-entier (ici 51V). Un recouvrement de signaux est donc souvent observé, limitant l'exploitation des données RMN. Un travail sur le découplage {51V} en RMN du 31P a donc été réalisé afin d'améliorer considérablement la résolution des spectres. Par la suite, en s'intéressant à la fois aux déplacements chimiques isotropes, à l'exploitation de séquences RMN 2D (jusque-là très peu utilisées) et à l'information structurale contenue dans le tenseur de déplacement chimique anisotrope (CSA) en 31P, il a été possible d'obtenir des renseignements sur la géométrie locale des groupements tétraédriques peu distordus PO4 [7]. Ainsi, en combinant ces résultats avec les calculs GIPAW de paramètres RMN dans une approche « NMR crystallography », nous avons pu contribuer à l'édition spectrale des spectres RMN, à l'analyse des informations structurales locales extraites pour les composés VOPO4, et au test des modèles ou hypothèses structurales apportés par la diffraction.
Par ailleurs, un suivi operando de l'oxydation de (VO)2P2O7 dans des conditions proches des conditions catalytiques (air + butane) a été réalisé en diffraction des rayons X sur la ligne CRISTAL du synchrotron SOLEIL. Cette expérience a été novatrice dans le cadre de cette classe de matériaux puisqu'elle a permis, pour la première fois, de montrer le rôle prépondérant d'un polymorphe VOPO4 spécifique dans l'activité catalytique. La métastabilité et le désordre dynamique sur la position du vanadium dans les octaèdres VO6 déformés de cette phase, démontré par RMN du solide du 51V en température, en condition statique, pourrait être à l'origine de la bonne activité catalytique décrite pour ce polymorphe. Nous avons en outre réalisé un suivi in situ et quantitatif en DRX du refroidissement de cette phase active en catalyse, en prenant en compte la fraction amorphe, ce qui a permis de comprendre le devenir de cette phase dans le catalyseur industriel post mortem, à la fin du processus catalytique.
Enfin, une combinaison in situ et ex situ de la RMN du solide et de la DRX dans la caractérisation des polymorphes γ- et δ-VOPO4 a permis de mettre en évidence une réorganisation de l'ordre local, et l'existence de contradictions avec les données publiées [8]. Ces informations, pressenties notamment par des informations microstructurales extraites de diagrammes de DRX par les poudres et de traînées de diffusion diffuse en DRX sur monocristal, ont pu être renforcées au travers d'expériences RMN 2D. Enfin, dans le cadre de l'étude du polymorphe δ-VOPO4, le croisement entre des calculs DFT de paramètres RMN, des expériences in situ en DRX par les poudres et en RMN du solide du 31P a permis de montrer une réorganisation de l'ordre local due à la température. Nous avons pu ainsi expliquer, entre autre, les signaux RMN excédentaires lorsque ce polymorphe est synthétisé ex situ, non expliqués directement par DRX.
Références
[1] Conte M., Budroni G., Bartley J. K., Taylor S. H., Carley A. F., Schmidt A., Murphy D. M., Girgsdies F., Ressler T., Schlögl R., Hutchings G. J., Science, 2006, 313, 1270-1273
[2] Dummer N.F., Bartley J.K. and Hutchings G.J., Advances in Catalysis, 2011, 189-247
[3] Duvauchelle N., Bordes E., Catalysis Letters, 1999, 57 (1-2), pp 81-88
[4] Eichelbaum M., Hävecker M., Heine C., Karpov A., Dobner C.-K., Rosowski F., Trunschke A., Schlögl R., Angewandte Chemie International Edition, 2012, 51 (25), pp 6246-6250
[5] Lapina, O. B. et al., J. of Mol. Catal. A : Chem., 162, 2000, 1, pp 381-390
[6] Ben Abdelouahab, F. et al., Journal of Catalysis, 134, 1992, pp 151-167
[7] Orr R. M., Duer M. J., Ashbrook S. E., Journal of Magnetic Resonance, 174, 2005, 2, pp 301-309
[8] Harlow R.L., Li Z.G., Herron N., Horowitz H.S., McCarron E.M., Richardson J.W. jr., Toby B.H. ICSD N°415213, 2005
Le prix Physique (Virginie Théry): Mon travail de thèse au laboratoire IRCer de Limoges a porté sur l'élaboration et la caractérisation structurale et physique de films minces à base de matériaux "intelligents", dont les propriétés électroniques peuvent être modifiées sous l'action d'une excitation externe (optique, électrique, thermique). Parmi cette catégorie de matériaux, mes études se sont portées sur le dioxyde de vanadium VO2, dont la transition métal-isolant à 68°C est également accompagnée d'une transition structurale entre une phase monoclinique (isolante) et une phase rutile (métallique) [1]. L'objectif principal de ma thèse a consisté à déterminer le rôle de la microstructure des films, et en particulier des contraintes épitaxiales, sur les transitions structurales et électriques. Pour cela, des films minces de VO2 de plusieurs épaisseurs ont été déposés par évaporation par faisceau d'électrons et par ablation laser sur différents substrats (Al2O3 (0001), TiO2 (001) et TiO2 (111)) [2].
Mon travail de thèse au laboratoire IRCer de Limoges a porté sur l'élaboration et la caractérisation structurale et physique de films minces à base de matériaux "intelligents", dont les propriétés électroniques peuvent être modifiées sous l'action d'une excitation externe (optique, électrique, thermique). Parmi cette catégorie de matériaux, mes études se sont portées sur le dioxyde de vanadium VO2, dont la transition métal-isolant à 68°C est également accompagnée d'une transition structurale entre une phase monoclinique (isolante) et une phase rutile (métallique) [1]. L'objectif principal de ma thèse a consisté à déterminer le rôle de la microstructure des films, et en particulier des contraintes épitaxiales, sur les transitions structurales et électriques. Pour cela, des films minces de VO2 de plusieurs épaisseurs ont été déposés par évaporation par faisceau d'électrons et par ablation laser sur différents substrats (Al2O3 (0001), TiO2 (001) et TiO2 (111)) [2].
Dans le cas de la croissance sur saphir, le fort désaccord de réseau promeut un mécanisme de croissance par coïncidence de domaines ce qui conduit, paradoxalement, à l'obtention de dépôts d'excellente qualité cristalline, notamment via la formation de dislocations géométriques dont les champs de distorsions sont fortement confinés à l'interface. Dans ce cas, la croissance se produit dans un état relaxé et l'essentiel des déformations est stocké au refroidissement du fait du désaccord de coefficients de dilatation. Une déformation en tension est alors appliquée le long de l'axe c de VO2 ce qui conduit à une ouverture du gap et à une stabilisation de la phase isolante monoclinique. L'étude de la transition structurale a permis de mettre en évidence l'apparition d'une phase secondaire au cours de la transition, dont l'analyse par spectroscopie Raman suggère qu'il s'agit de la phase monoclinique M2 de VO2 [3].
La croissance sur TiO2 (001) est caractérisée par un désaccord de réseau plus faible (1%) avec une épaisseur critique (critère de Matthews et Blakeslee) de l'ordre de 4 nm, épaisseur à partir de laquelle des dislocations de type [101](101) glissent jusqu'à l'interface en vue de relaxer l'énergie élastique. En plus de ces dislocations, une relaxation par formation de lacunes d'oxygènes (facilitée par les degrés d'oxydation multiples du vanadium) est également observée. L'analyse de cartographies de l'espace réciproque permet de conclure que l'évolution des transitions électriques et structurales résulte de la compétition entre les déformations épitaxiales, les déformations d'origines thermiques et la présence de lacunes d'oxygène à l'interface et leurs influences respectives sur les orbitales moléculaires de VO2 [4]. 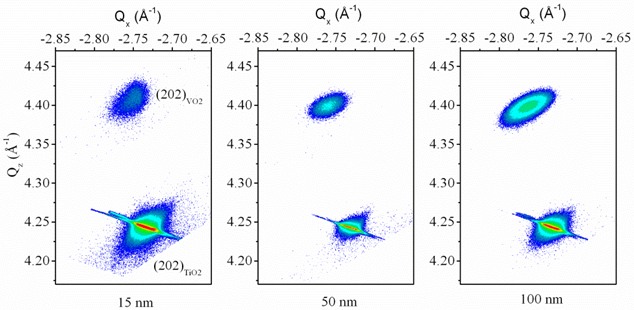
Figure : Cartographie de l'espace réciproque de la réflexion (202)TiO2 et de la réflexion (202)VO2 obtenue pour des films minces de 15, 50 et 100 nm d'épaisseur de VO2 déposés sur un substrat de TiO2 orienté (001).
Enfin, la croissance des couches minces sur TiO2 (111) est également caractérisée par un désaccord de réseau de l'ordre du pourcent. Les études structurales et électriques montrent une diminution de la température de transition lorsque l'épaisseur des couches minces augmente. Comme pour l'orientation (001) de TiO2, l'analyse de l'origine des déformations au sein des films minces a mis en évidence le phénomène de relaxation de la couche de VO2 par formation de lacunes d'oxygène au sein du réseau au cours de la croissance. La présence des lacunes d'oxygène permet ainsi d'expliquer les valeurs de température de transition inférieures à la valeur de VO2 massif (68°C).
Références :
[1] F.J. Morin, Phys. Rev. Lett., 3 :34, 1959.
[2] V. Théry, A. Boulle, A. Crunteanu, J. C. Orlianges, A. Beaumont, R. Mayet, A. Mennai, F. Cosset, A. Bessaudou and M. Fabert. J. Appl. Phys., 121 :055303, 2017.
[3] V. Théry, A. Boulle, A. Crunteanu, J. C. Orlianges, A. Beaumont, R. Mayet, A. Mennai, F. Cosset, A. Bessaudou and M. Fabert. Phys. Rev. B, 93 :184106, 2016.
[4] V. Théry, A. Boulle, A. Crunteanu, J. -C. Orlianges, Appl. Phys. Lett., 111 :251902, 2017.