
L'AFC félicite ses 3 lauréats du prix de thèse 2013. Les lauréats présenteront leurs travaux en cours de l'année 2014 lors d'un évenement à définir.
Le prix AFC - Mention Biologie a été attribué à Romain Talon, Institut de Biologie Structurale Jean-Pierre Ebel de Grenoble, thèse soutenue en juin 2012 sous la direction de Richard Kahn et Eric Girard. Titre - Développement de nouveaux outils pour la détermination de la structure de macromolécules biologiques par diffraction : application aux protéines membranaires et aux grands complexes protéiques.
Le prix AFC - Mention Chimie a été attribué à Nicolas Leblanc, Laboratoire MOLTECH Anjou, Université d’Angers, thèse soutenue en décembre 2011 sous la direction du Professeur Nicolas Mercier. Titre - Halobismuthates et haloantimonates de viologène : ferroélectricité et transfert de charge photo-induit.
Le prix AFC - Mention Physique a été attribué à Céline Mariette, Institut de Physique de Rennes, thèse soutenue en juin 2013 sous la direction de Bertrand Toudic et Philippe Rabiller. Titre - Brisures de symétrie dans des superespaces cristallographiques : aspects structuraux et dynamiques.
Le prix Biologie (Romain Talon):

Ces travaux de recherche s’inscrivent dans le cadre des développements méthodologiques pour la biocristallographie. Ils ont consisté en l’amélioration, à l’élaboration et à l’utilisation de complexes de lanthanide en tant qu’outils permettant de faciliter la détermination de structures de macromolécules biologiques par diffraction des rayons X. Ces complexes sont constitués d’un ion lanthanide chélaté par un ligand. Ce ligand peut, d’une part, assurer une liaison non-covalente avec les surfaces de macromolécules biologiques et d’autre part, rendre le complexe luminescent dans le spectre visible, ce qui permet de révéler aisément sa fixation dans les cristaux de protéines avant l’étape de diffraction. Une fois les complexes introduits dans les cristaux par co-cristallisation ou par trempage, les atomes de lanthanide constituent quant à eux une sous-structure qui, déterminée par les méthodes de phasage de novo courantes (SAD, MAD, SIRAS, MIRAS), permet de résoudre la structure de la macromolécule d’intérêt, même si celle-ci est de grande taille. En effet, au travers de huit fructueuses collaborations et trente structures résolues par phasage expérimental, ces complexes ont notamment permis d’obtenir par la méthode SAD la structure de l’aminopeptidase homo-dodécamérique TET1 de 444 kDa à 4 Å de résolution (Talon et al., 2011). Aussi, les structures de trois nouvelles enzymes homo-tétramériques de la famille des malate déshydrogénases ont été déterminées à très haute résolution. D’une part, en utilisant ces enzymes en tant que bibliothèque de fonctions chimiques, nous avons pu mettre en place une nouvelle approche méthodologique permettant une compréhension plus fine des modes d’interaction des complexes de lanthanide.
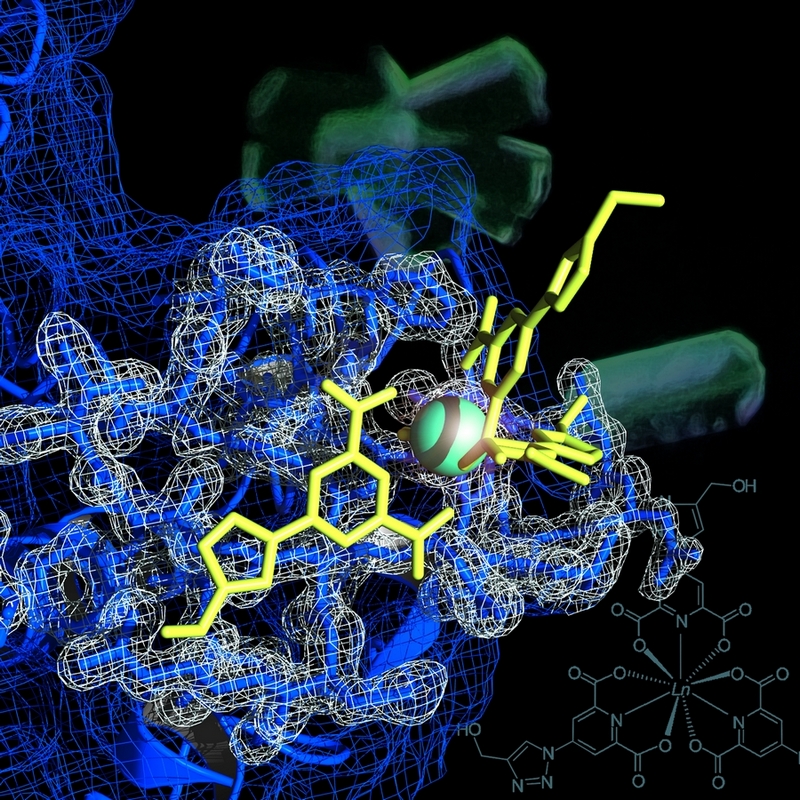
D’autre part, ceci a aussi donné lieu à de nombreux résultats annexes qui apportent de nouveaux éclaircissements sur l’adaptation halophile (Coquelle et al., 2010, Talon et al., 2013, soumis pour publication). Enfin, une nouvelle famille de complexes luminescents, modulables dans leur structure et présentant une plus grande affinité pour les surfaces des protéines modèles usuelles, a été élaborée et caractérisée (Talon et al., 2012, dépôt de brevet n°1255216). Ces précieux résultats scientifiques n’ont pu être obtenus que grâce à une ambiance de travail aussi joviale que studieuse, donc particulièrement exaltante, sous l’égide du Dr. Eric Girard et du Dr. Richard Kahn.
La thèse - sur le serveur TEL
R. Talon, L. Nauton, J. L. Canet, R. Kahn, E. Girard, A. Gautier, ChemComm 48, 11886-11888 (2012)
R. Talon, R. Kahn, M. A. Durá, O. Maury, F. M. D. Vellieux, B. Franzetti, E. Girard, J. Synchr. Rad. 18, 74-78 (2011)
N. Coquelle, R. Talon, D.H. Juers, E. Girard, R. Kahn, D. Madern, J. Mol. Biol. 404, 493-505 (2010)
Le prix Chimie (Nicolas Leblanc):

Ce travail de thèse s’inscrit dans la thématique très large des hybrides organiques-inorganiques, dont le but est d’obtenir des matériaux combinant à la fois les propriétés uniques d’une partie inorganique (semi-conductivité, magnétisme, luminescence, propriétés optiques..), aux propriétés uniques d’une partie organique (flexibilité, polarisabilité, matériau filmogène...). Dans notre étude nous avons combiné des anions inorganiques, provenant des éléments Bismuth (Bi3+), Antimoine (Sb3+) et halogénure (Cl-, Br-, I-), à une molécule organique bien connue, le Méthylviologène. Cette association nouvelle nous a permis d’identifier de nouveaux matériaux présentant d’intéressantes propriétés : -Dans une première étude, nous avons mis à jour une nouvelle famille de composés présentant des propriétés ferroélectriques remarquables, phénomène électrique (polarisation commutable par un champ électrique) équivalent à celui déjà rencontré dans les matériaux aimants (ferromagnétiques), et également à la base de nombreuses technologies, notamment dans le stockage de données et l’électronique (Disques Durs, Mémoires FeRam…). -Dans une deuxième étude, notre attention s’est portée sur une série de matériaux présentant d’intéressantes propriétés photochromes (changement de couleur du matériau sous l’action de la lumière et ce de façon réversible). Ce changement de couleur étant lié à la formation de charges stabilisées à l’intérieur du composé, il était donc important d’en comprendre les mécanismes afin de pouvoir par la suite utiliser potentiellement ces matériaux dans des applications comme le stockage d’énergie, ou encore dans le photovoltaïque, lors du relargage des charges dans un circuit électrique.

Le prix Physique (Céline Mariette):

La symétrie de translation qui a longtemps caractérisé le cristal est à la base de l’analyse de la physique du solide. Ces dernières décennies, la définition du cristal a changé incluant désormais des matériaux qui possèdent un ordre à grande distance, mais sans la symétrie de translation à trois dimensions. Ces composés dits « apériodiques » sont décrits dans le cadre de la cristallographie de superespace, espace de dimension supérieure à trois dans lequel la périodicité est retrouvée [1]. Un exemple fameux est donné par les quasicristaux, une découverte récompensée par le prix Nobel de Chimie 2011 [2]. Il existe d’autres types de cristaux apériodiques, dont les cristaux composites d’inclusion non stochéométriques. Durant cette thèse, nous nous sommes particulièrement intéressés à une famille prototype de composés supramoléculaires organiques, les n-alcanes/urée. Ces cristaux sont constitués d’un réseau d’urée qui forme des canaux dans lesquels sont imbriquées les chaines linéaires d'alcane. A température ambiante, l’urée a une structure en nid d’abeille de symétrie hexagonale, et les molécules d’alcanes présentent un désordre de rotation autour de leur axe. Suivant la direction des canaux, les périodicités des deux sous-réseaux sont généralement dans un rapport irrationnel. On peut aisément obtenir de tels cristaux d’inclusion avec des molécules d’alcane CnH2n+2 pour n plus grand que sept (heptane), ce qui fait de cette famille une plateforme modèle pour l’étude de la physique des composites apériodiques. Du fait des nombreux degrés de libertés offerts par l’apériodicité, ces composés présentent des transitions de phase structurales très originales [3].
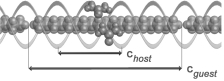
Des études systématiques par diffraction de rayons X sur sources synchrotron révèlent de multiples solutions structurales impliquant des changements ou non de la dimension du groupe de superespace. Une fois la caractérisation du paramètre d’ordre et de la brisure de symétrie faite; nous avons pour la première fois pu étudier des phénomènes prétransitionnels critiques dans des superespaces avec en l’occurrence une augmentation de la dimension du superespace [4]. La diffusion cohérente de neutrons et la diffusion inélastique de rayons X nous ont permis une analyse dynamique des différentes excitations dans ces matériaux, en particulier les modes d’excitations spécifiques aux cristaux apériodiques, c’est à dire selon les directions internes du superespace (les phasons).
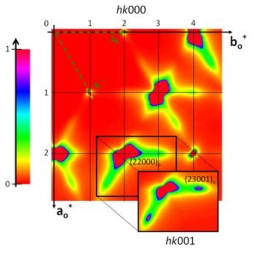
Les composés d’inclusion avec des molécules invitées courtes (n variant de 7 à 13) présentent en outre à température ambiante des phases « quasi-liquides » unidimensionnelles. Ce désordre dynamique le long de la direction d'incommensurabilité de ces matériaux génère à basse température des solutions structurales nouvelles (composite monoclinique intermodulé, ancrages commensurables), exploitant des degrés de glissement à coût nul prédits dans de tels matériaux apériodiques [5].
Références:
[1] T. Janssen, G. Chapuis, M. de Boissieu, Aperiodic Crystals: From Modulated Phases to Quasicrystals, (Oxford Univ. Press, Oxford, 2007).
[2] D. Schechtman, I. Blech, D. Gratias and J. W. Cahn, Phys. Rev. Lett. 53, 1951–1953 (1984)
[3] B. Toudic, P. Garcia, C. Odin, et al., Science 319 (2008), 69-72.
[4] C. Mariette, L. Guérin, P. Rabiller, et al., Phys. Rev. B87, 104101 (2013)
[5] C. Mariette, M. Huard, P. Rabiller, et al., J. Chem. Phys.136, 104507 (2012)
L'AFC félicite ses 3 lauréats du prix de thèse 2014. Les lauréats ont présenté leurs travaux lors du colloque Rayons-X et Matière du 1er au 4 décembre 2015 à Grenoble.
Le prix AFC - Mention Biologie a été attribué à Ali Flayhan, Institut de Biologie Structurale à Grenoble, thèse soutenue en juin 2012 sous la direction de Cécile Breyton. Titre: Reconnaissance phage – bactérie dans le système phage T5 – E. coli. Caractérisation biochimique et structurale du complexe FhuApb5 et de la protéine caudale pb9.
Le prix AFC - Mention Physique a été attribué à Mohamed Salah Amara, Laboratoire de Physique des Solides d'Orsay, thèse soutenue en décembre 2014 sous la direction de Pascale Launois et la co-direction d'Antoine Thill au LIONS, CEA-Saclay. Titre - Nanotubes d'imogolite et propriétés de l'eau confinée : organisation, structure et dynamique
Le prix AFC - Mention Chimie a été attribué à Arnaud Grosjean, Institut de Chimie de la Matière Condensée de Bordeaux, thèse soutenue en décembre 2013 sous la direction de Philippe Guionneau. Titre - Matériaux polymériques 1D à transition de spin: investigations structurales multi-échelles.
Le prix Biologie (Ali Flayhan):
 Au cours de cette thèse, j’ai abordé les premières étapes de l’infection d’une bactérie par un bactériophage dans le système E. coli - phage T5. Mes travaux de recherche se sont principalement focalisés sur la caractérisation biochimique et structurale du complexe formé entre pb5, la RBP (Receptor Binding Protein) de T5 et son récepteur FhuA, un transporteur de fer-ferrichrome, de la membrane externe d’E. coli. Cette interaction induit très probablement des changements de conformation, transmis le long de la queue jusqu'à la capside. Ils provoquent l'ouverture de la capside et l'éjection de l'ADN viral dans le cytoplasme bactérien, via un canal formé par le virus qui perfore la paroi bactérienne. J’ai montré que la formation du complexe n’induit pas de réarrangements majeurs des structures de pb5 et/ou de FhuA. Seuls des changements de conformation subtils, au niveau de structures secondaires, ont été décelés et attribués à pb5. Ces changements de conformation seraient à l’origine de la transmission du signal au reste du phage. Par ailleurs, des cristaux 3D de qualité moyenne (diffraction rayons X 8Å) et 2D de qualité supérieure (diffraction électronique 3Å) ont été obtenus. Des études de diffusion de neutrons et de rayons X aux petits angles ont permis d’obtenir un premier modèle tridimensionnel de pb5 seule et au sein du complexe. Ces modèles sont en accord avec la structure à basse résolution de pb5 et du complexe, obtenues par microscopie électronique sur particules isolées, et montrent que l’interface de liaison entre les deux protéines couvre toute la section extracellulaire de FhuA. De plus, pb5 se lie à FhuA par l’une de ses extrémités de telle manière que son grand axe et l’axe du tonneau de FhuA soient alignés (Fig. 1). Par ailleurs, contrairement aux différentes RBP décrites jusqu’à présent, pb5 semble composée d’un domaine unique et est présente en une seule copie au bout distal de la fibre droite de T5.
Au cours de cette thèse, j’ai abordé les premières étapes de l’infection d’une bactérie par un bactériophage dans le système E. coli - phage T5. Mes travaux de recherche se sont principalement focalisés sur la caractérisation biochimique et structurale du complexe formé entre pb5, la RBP (Receptor Binding Protein) de T5 et son récepteur FhuA, un transporteur de fer-ferrichrome, de la membrane externe d’E. coli. Cette interaction induit très probablement des changements de conformation, transmis le long de la queue jusqu'à la capside. Ils provoquent l'ouverture de la capside et l'éjection de l'ADN viral dans le cytoplasme bactérien, via un canal formé par le virus qui perfore la paroi bactérienne. J’ai montré que la formation du complexe n’induit pas de réarrangements majeurs des structures de pb5 et/ou de FhuA. Seuls des changements de conformation subtils, au niveau de structures secondaires, ont été décelés et attribués à pb5. Ces changements de conformation seraient à l’origine de la transmission du signal au reste du phage. Par ailleurs, des cristaux 3D de qualité moyenne (diffraction rayons X 8Å) et 2D de qualité supérieure (diffraction électronique 3Å) ont été obtenus. Des études de diffusion de neutrons et de rayons X aux petits angles ont permis d’obtenir un premier modèle tridimensionnel de pb5 seule et au sein du complexe. Ces modèles sont en accord avec la structure à basse résolution de pb5 et du complexe, obtenues par microscopie électronique sur particules isolées, et montrent que l’interface de liaison entre les deux protéines couvre toute la section extracellulaire de FhuA. De plus, pb5 se lie à FhuA par l’une de ses extrémités de telle manière que son grand axe et l’axe du tonneau de FhuA soient alignés (Fig. 1). Par ailleurs, contrairement aux différentes RBP décrites jusqu’à présent, pb5 semble composée d’un domaine unique et est présente en une seule copie au bout distal de la fibre droite de T5.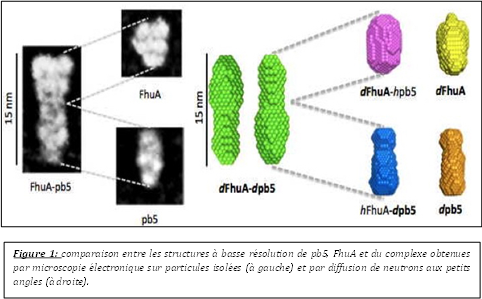 En parallèle de l’étude du complexe FhuA-pb5 et dans le cadre d’un projet visant à localiser et déterminer les structures de l’ensemble des protéines caudales de T5, j'ai purifié, cristallisé et résolu la structure de pb9, une des protéines de la queue de T5 composée de deux domaines A et B (Fig. 2). Le phasage a été particulièrement laborieux car aucune structure homologue n’était disponible et les données anomales étaient de très mauvaise qualité. En effet, je n’ai pu avoir des données anomales qu’avec les complexes lanthanides, tous mes efforts, pour obtenir des meilleures données anomales à partir des trempages avec les sels d'atomes lourds classiques, ayant échoués. Nous avons localisé pb9 par immunolocalisation, en haut de la partie conique de la queue de T5. La structure nous a permis d’identifier pb9 comme étant la protéine distale de la queue (Dit) de T5, elle forme un anneau hexamérique qui relie la partie conique au tube central de la queue. Ce motif structural a été déjà observé chez les siphophages (phages à longue queue flexible non contractile) infectant des bactéries à Gram+. Pb9 est la première Dit décrite chez les siphophages infectant des bactéries à Gram-.
En parallèle de l’étude du complexe FhuA-pb5 et dans le cadre d’un projet visant à localiser et déterminer les structures de l’ensemble des protéines caudales de T5, j'ai purifié, cristallisé et résolu la structure de pb9, une des protéines de la queue de T5 composée de deux domaines A et B (Fig. 2). Le phasage a été particulièrement laborieux car aucune structure homologue n’était disponible et les données anomales étaient de très mauvaise qualité. En effet, je n’ai pu avoir des données anomales qu’avec les complexes lanthanides, tous mes efforts, pour obtenir des meilleures données anomales à partir des trempages avec les sels d'atomes lourds classiques, ayant échoués. Nous avons localisé pb9 par immunolocalisation, en haut de la partie conique de la queue de T5. La structure nous a permis d’identifier pb9 comme étant la protéine distale de la queue (Dit) de T5, elle forme un anneau hexamérique qui relie la partie conique au tube central de la queue. Ce motif structural a été déjà observé chez les siphophages (phages à longue queue flexible non contractile) infectant des bactéries à Gram+. Pb9 est la première Dit décrite chez les siphophages infectant des bactéries à Gram-. 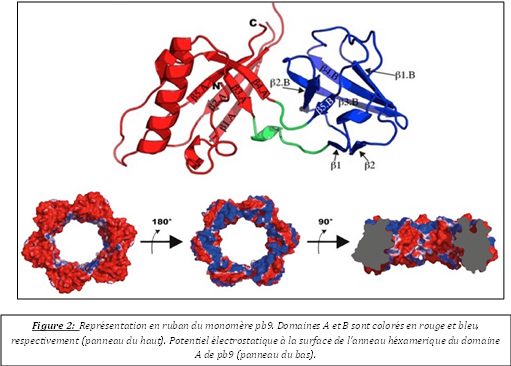 Nous proposons que ce motif soit conservé parmi tous les siphophages.
Nous proposons que ce motif soit conservé parmi tous les siphophages.Références:
- M. Noirclerc-Savoye, A. Flayhan, C. Pereira, B. Galet, P. Gans, C. Ebel, C. Breyton, Protein Expr Purif., 109C, 70-78 (2015).
- A. Flayhan, F. M. Vellieux, R. Lurz, O. Maury, C. Contreras-Martel, E. Girard, P. Boulanger, C. Breyton, J. Virol., 88(2), 820-828 (2014).
- Y. Zivanovic, F. Confalonieri, L. Ponchon, R. Lurz, M. Chami, A. Flayhan, M. Renouard, A. Huet, P. Decottignies, A. R. Davidson, C. Breyton, P. Boulanger, J. Virol., 88(2), 1162-1174 (2014).
- C. Breyton, A. Flayhan, F. Gabel, M. Lethier, G. Durand, P. Boulanger, M. Chami, C. Ebel, J. Biol. Chem., 288(42), 30763-30772 (2013).
- C. Breyton, F. Gabel, M. Lethier, A. Flayhan, G. Durand, J.M. Jault, C. Juillan-Binard, L. Imbert, M. Moulin, S. Ravaud, M. Härtlein, C. Ebel, Eur. Phys. J. E Soft Matter, 36(7), 71 (2013).
- A. Flayhan, F. Wien, M. Paternostre, P. Boulanger, C. Breyton, Biochimie, 94(9), 1982-1989 (2012).
Le prix Physique (Mohamed Salah Amara): Ce travail porte sur les propriétés structurales de nano-containers modèles ainsi que sur les propriétés structurales et dynamique de l’eau nano-confinée. Il s’inscrit dans les domaines de la nanophysique et de la nanofluidique.
Ce travail porte sur les propriétés structurales de nano-containers modèles ainsi que sur les propriétés structurales et dynamique de l’eau nano-confinée. Il s’inscrit dans les domaines de la nanophysique et de la nanofluidique.
Sa morphologie tubulaire unidimensionnelle et son diamètre de l’ordre du nanomètre font de l'imogolite, nanotube d'aluminosilicate (AlSi) ou aluminogermanate (AlGe) de formule générique HOSixGe1-xO3Al2(OH)3, un nano-container idéal. Mon travail de thèse s'est articulé autour de trois axes principaux : la synthèse des nanotubes [1,2,3], l'étude de leur structure et l'étude de fluides confinés, en particulier l'eau. Ce sont les deux dernières parties de ce travail, menées au Laboratoire de Physique d’Orsay ainsi qu’à l’ILL à Grenoble, qui sont récompensées par le prix de thèse de l’AFC, mention Physique.
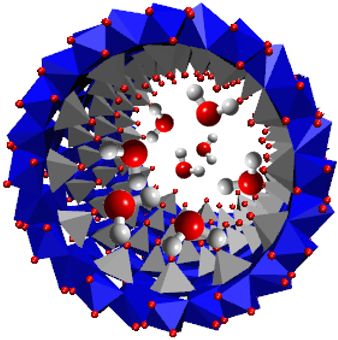 La détermination de la structure atomique de différents nanotubes d'imogolite, naturels et synthétiques, à base de silicium ou de germanium, mono- et double-parois a été réalisée grâce à des expériences de diffusion des rayons X aux petits et aux grands angles sur des suspensions de nanotubes et à la simulation des diagrammes DRX obtenus, sur la base d'un développement sur les fonctions de Bessel cylindriques [4]. Le point original de l'étude mise en place est l'élaboration d'une stratégie qui nous a permis de rendre compte à la fois des données DRX aux petits et aux grands angles (Amara et al., en préparation). Elle est basée sur la minimisation des distances interatomiques et des angles entre les liaisons au sein des tétraèdres SiO4 (GeO4) et des octaèdres AlO6 qui constituent une paroi du nanotube. Cette étude exhaustive a permis d'obtenir le nombre d'unités structurales par circonférence du nanotube ainsi que les coordonnées atomiques. Nous avons ainsi pu, pour la première fois, élucider la structure exacte des nanotubes d’aluminosilicates naturels -découverts en 1962- et synthétiques -obtenus dès 1977- et des nanotubes mono-parois d’aluminogermanates. Pour les nanotubes d'imogolite AlGe double-parois, nous avons de plus mis en évidence, pour la première fois aussi, l'existence de deux périodes incommensurables sur les deux parois interne et externe du tube.
La détermination de la structure atomique de différents nanotubes d'imogolite, naturels et synthétiques, à base de silicium ou de germanium, mono- et double-parois a été réalisée grâce à des expériences de diffusion des rayons X aux petits et aux grands angles sur des suspensions de nanotubes et à la simulation des diagrammes DRX obtenus, sur la base d'un développement sur les fonctions de Bessel cylindriques [4]. Le point original de l'étude mise en place est l'élaboration d'une stratégie qui nous a permis de rendre compte à la fois des données DRX aux petits et aux grands angles (Amara et al., en préparation). Elle est basée sur la minimisation des distances interatomiques et des angles entre les liaisons au sein des tétraèdres SiO4 (GeO4) et des octaèdres AlO6 qui constituent une paroi du nanotube. Cette étude exhaustive a permis d'obtenir le nombre d'unités structurales par circonférence du nanotube ainsi que les coordonnées atomiques. Nous avons ainsi pu, pour la première fois, élucider la structure exacte des nanotubes d’aluminosilicates naturels -découverts en 1962- et synthétiques -obtenus dès 1977- et des nanotubes mono-parois d’aluminogermanates. Pour les nanotubes d'imogolite AlGe double-parois, nous avons de plus mis en évidence, pour la première fois aussi, l'existence de deux périodes incommensurables sur les deux parois interne et externe du tube.
L'étude structurale des imogolites nous a de plus amené à étudier leur déformation éventuelle. Nous avons d'abord montré comment contrôler l'assemblage des imogolites en phase solide, via le contrôle de la salinité de la suspension initiale. Nous avons alors pu étudier la déformation des imogolites AlGe mono-parois selon qu’ils sont organisés en petits (3-4 tubes) ou en grands (~60 tubes) fagots. L'analyse détaillée des mesures DRX effectuées sur ces deux sortes d'échantillons en poudre montre que les tubes acquièrent une forme elliptique dans le premier cas, en accord avec les prédictions théoriques, et qu'ils adoptent une forme hexagonale quand ils sont agglomérés en gros fagots selon un réseau hexagonal bidimensionnel. La combinaison d'un modèle analytique adapté et des mesures DRX montrent pour la première fois l'hexagonalisation des imogolites, introduisant un nouveau membre dans la famille déjà riche des imogolites [5].
Le nano-confinement à l'échelle moléculaire induit de nouvelles propriétés des molécules confinées. Les imogolites, nano-canaux hydrophiles avec un diamètre de l'ordre du nanomètre, constituent un système unique pour étudier le nano-confinement de l'eau. Nous nous sommes donc intéressés à l'étude de sa structure et de sa dynamique vibrationnelle. L'analyse des expériences de DRX in situ en fonction de la température montre l'existence de trois domaines différents en température, correspondant au départ des molécules d'eau à l'extérieur des nanotubes, puis de celles au centre des nanotubes (eau dite "confinée") et enfin des molécules d'eau en interaction avec la paroi interne (eau dite "liée"). Les deux dernières sortes d'eau présentent une dynamique vibrationnelle très différente : la diffusion inélastique des neutrons montre la signature d'une eau "amorphe" pour l'eau confinée, et la disparition totale des modes acoustiques dans le cas de l'eau liée avec l'apparition d'un nouveau mode autour de 13 meV, que nous attribuons à l'élongation des liaisons hydrogène avec la surface. Les modes de librations intermoléculaires présentent un amollissement par rapport au cas de l'eau en volume, plus important pour l'eau liée que pour l'eau confinée, en accord avec la diminution du nombre de liaisons hydrogène pour l'eau confinée et la diminution des degrés de libertés de libration pour l'eau liée. En résumé, notre étude, la première à avoir été réalisée sur ce système, a démontré l'existence de deux sortes d'eau dans les nanotubes, avec des propriétés dynamiques différentes (Amara et al., en préparation).
[1] M.S. Amara, E. Paineau, M. Bacia-Verloop, M.-E. M. Krapf, P. Davidson, L. Belloni, C. Levard, J. Rose, P. Launois, A. Thill. Chem. Commun. 49, 11284 (2013).
[2] A. Thill and M.S. Amara. Method for producing germanium imogolite nanotubes, patent WO 2014/147579 A1 (2014).
[3] M.S. Amara, E. Paineau, S. Rouzière, B. Guiose, M.-E. M. Krapf, O. Taché, P. Launois, A. Thill. Chem. Mater. 27, 1488 (2015).
[4] M.S. Amara, C. Mariette, E. Paineau, S. Rouzière, D. Petermann, M. Kociak, P. Launois. Reflets Phys. N°44-45, 34 (2015).
[5] M.S. Amara, S. Rouzière, E. Paineau, M. Bacia-Verloop, A. Thill, P. Launois. J. Phys. Chem. C 118, 9299 (2014).
Le prix Chimie (Arnaud Grosjean):

[2] G. Aromi et al., Coordination Chemistry Reviews 255 (2011) 485-546
[3] O. Roubeau, Chem. Eur. J, 18 (2012) 15230-15244
[4] P. Guionneau et al., Top. Curr. Chem. 234 (2004) 97-128
[5] M. A. Halcrow, Chem. Soc. Rev., 40 (2011) 4119-4142
[6] J. G. Haasnoot et al., Z. Naturforsch, 32b(1977) 1421
[7] A. Urakawa et al., J. Phys. Chem. C, 115 (2011) 1323-1329
[8] A. Michalowicz et al., Chem. Mater., 7 (1995) 1833-1842
[9] A. Grosjean et al., Chem. Commun., 47 (2011) 12382-12384
[10] A. Grosjean et al., Eur. J. Inorg. Chem. (2013) 796-802
L'AFC félicite ses 3 lauréats du prix de thèse 2016. Les lauréats ont présenté leurs travaux lors du colloque de l'Association Française de Cristallographie du 4 au 7 juillet 2016 à Marseille.
Le prix AFC - Mention Biologie a été attribué à Guillaume Gaullier, Institut de Biologie Intégrative de la Cellule (I2BC) à Gif-sur-Yvette, thèse soutenue le 22 septembre 2015 sous la direction de Marie-Hélène Le Du. Titre: Étude structurale de l’assemblage du complexe télomérique humain TRF2/RAP1.
Le prix AFC - Mention Chimie a été attribué à Morgane Poupon, Laboratoire de Cristallographie et Sciences des Matériaux à Caen, thèse soutenue le 9 novembre 2015 sous la direction de Nicolas Barrier et Olivier Pérez. Titre - Synthèse d’oxydes à base d’éléments à paire libre (SeIV et TeIV) dans le but d’obtenir des structures cristallines non-centrosymétriques originales.
Le prix AFC - Mention Physique a été attribué à Maxime Dupraz, Laboratoire de Science et Ingénierie des Matériaux et Procédés (SIMAP) à Grenoble, thèse soutenue le 17 novembre 2015 sous la direction de Marc Verdier et Guillaume Beutier. Titre - Diffraction des rayons X cohérents appliquée à la physique du métal.
Le prix Biologie (Guillaume Gaullier):
 Effectuée au CEA de Saclay sous la direction de Marie-Hélène Le Du, ma thèse a porté sur l'étude structurale du complexe entre TRF2 (telomere repeat-binding factor 2) et RAP1 (repressor/activator protein 1), deux protéines télomériques humaines. Les télomères sont les extrémités des chromosomes des eucaryotes, ils sont constitués de répétitions d'un motif court riche en guanine et lié par des protéines spécifiques. L'intégrité des télomères est critique pour garantir la stabilité du génome. TRF2 et RAP1 sont les protéines télomériques les plus abondantes chez les mammifères (Takai et al, 2010), et sont essentielles pour établir un télomère fonctionnel : le retrait de TRF2 des télomères entraine en effet l'activation des voies de signalisation des dommages de l'ADN (Takai et al, 2003), et une fréquence élevée de fusions de chromosomes (voir par exemple Sarthy et al, 2009). Quand ma thèse a débuté, nous savions déjà que RAP1 s'associe à TRF2, par son domaine C-terminal et avec une affinité nanomolaire (Chen et al, 2011). Nous disposions aussi de structures tridimensionnelles de domaines isolés (Chen et al, 2011; Court et al, 2005; Fairall et al, 2001; Hanaoka et al, 2001), mais aucune étude structurale n'avait été réalisée sur les protéines entières ou leur complexe. Ce type d'étude représente un défi car aucune méthode expérimentale ne permet à elle seule de répondre à toutes les questions que soulèvent
Effectuée au CEA de Saclay sous la direction de Marie-Hélène Le Du, ma thèse a porté sur l'étude structurale du complexe entre TRF2 (telomere repeat-binding factor 2) et RAP1 (repressor/activator protein 1), deux protéines télomériques humaines. Les télomères sont les extrémités des chromosomes des eucaryotes, ils sont constitués de répétitions d'un motif court riche en guanine et lié par des protéines spécifiques. L'intégrité des télomères est critique pour garantir la stabilité du génome. TRF2 et RAP1 sont les protéines télomériques les plus abondantes chez les mammifères (Takai et al, 2010), et sont essentielles pour établir un télomère fonctionnel : le retrait de TRF2 des télomères entraine en effet l'activation des voies de signalisation des dommages de l'ADN (Takai et al, 2003), et une fréquence élevée de fusions de chromosomes (voir par exemple Sarthy et al, 2009). Quand ma thèse a débuté, nous savions déjà que RAP1 s'associe à TRF2, par son domaine C-terminal et avec une affinité nanomolaire (Chen et al, 2011). Nous disposions aussi de structures tridimensionnelles de domaines isolés (Chen et al, 2011; Court et al, 2005; Fairall et al, 2001; Hanaoka et al, 2001), mais aucune étude structurale n'avait été réalisée sur les protéines entières ou leur complexe. Ce type d'étude représente un défi car aucune méthode expérimentale ne permet à elle seule de répondre à toutes les questions que soulèvent 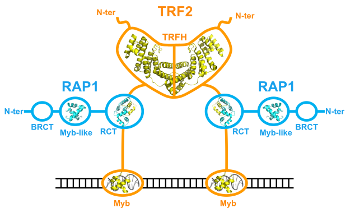 ce type de protéines multi-domaines. Mes travaux ont consisté à obtenir et intégrer des données issues de plusieurs méthodes expérimentales complémentaires : la cristallographie, mais aussi la diffusion des rayons X aux petits angles (SAXS), la microcalorimétrie, et des expériences d'empreinte protéique par acétylation des lysines. Les données de SAXS nous ont permis de confirmer que TRF2 est dimérique, RAP1 est monomérique, et le complexe TRF2/RAP1 contient deux monomères de RAP1 associés à un dimère de TRF2. Nos données suggèrent également un changement de conformation des protéines lors de l'assemblage du complexe. Une étude par microcalorimétrie comparant les propriétés d'interaction avec TRF2 de RAP1 entière et du domaine C-terminal isolé de RAP1 a permis de distinguer un signal détectable uniquement en présence de RAP1 entière, signifiant que la seule liaison du domaine C-terminal de RAP1 à TRF2 ne rend pas complètement compte de leur mode d'interaction.
ce type de protéines multi-domaines. Mes travaux ont consisté à obtenir et intégrer des données issues de plusieurs méthodes expérimentales complémentaires : la cristallographie, mais aussi la diffusion des rayons X aux petits angles (SAXS), la microcalorimétrie, et des expériences d'empreinte protéique par acétylation des lysines. Les données de SAXS nous ont permis de confirmer que TRF2 est dimérique, RAP1 est monomérique, et le complexe TRF2/RAP1 contient deux monomères de RAP1 associés à un dimère de TRF2. Nos données suggèrent également un changement de conformation des protéines lors de l'assemblage du complexe. Une étude par microcalorimétrie comparant les propriétés d'interaction avec TRF2 de RAP1 entière et du domaine C-terminal isolé de RAP1 a permis de distinguer un signal détectable uniquement en présence de RAP1 entière, signifiant que la seule liaison du domaine C-terminal de RAP1 à TRF2 ne rend pas complètement compte de leur mode d'interaction.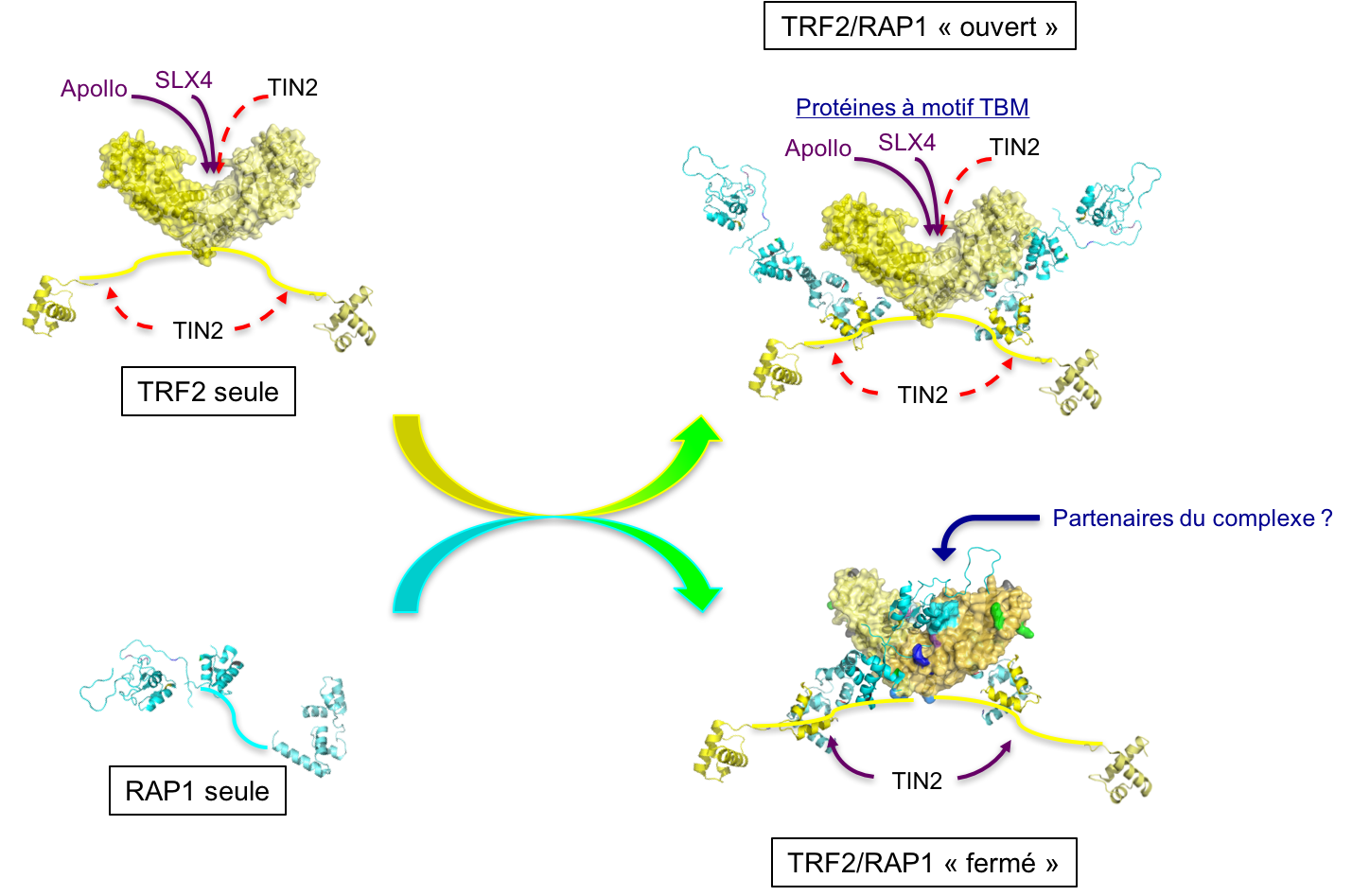 La région N-terminale de RAP1 contient un motif Y-x-L-x-P décrit dans la littérature comme un motif d'interaction avec le domaine TRFH (domaine de dimérisation) de TRF2 (Chen et al, 2008; Kim et al, 2009) et appelé TBM (TRFH-binding motif). La structure cristallographique du domaine TRFH de TRF2 en complexe avec un peptite de RAP1 contenant le motif Y-x-L-x-P montre qu'il s'agit bien d'un TBM, et la mutation en alanine de ces trois résidus dans RAP1 entière abolit le signal biphasique observé par microcalorimétrie, ce qui confirme l'implication de ce motif dans l'interaction entre les protéines entières. Finalement, les expériences d'empreinte protéique ont montré que l'interaction entre TRF2 et RAP1 engage de vastes surfaces des deux protéines, et que ces surfaces sont plus réduites dans le complexe contenant RAP1 mutée dans le TBM. L'ensemble de nos données est en faveur d'un modèle selon lequel RAP1 lie TRF2 avec une haute affinité par son domaine C-terminal, et peut selon les conditions occuper le site de liaison des motifs TBM dans le domaine TRFH de TRF2 (figure à droite). Ces résultats ont été publiés peu après la fin de ma thèse (Gaullier et al, 2016). Nous avons formulé deux hypothèses (non mutuellement exclusives) sur la fonction de ce mode d'interaction : la première propose qu'un des rôles de RAP1 pourrait être de réguler l'accès au site de liaison des motifs TBM par d'autres protéines contenant ces motifs, la seconde propose que la dimérisation de la région N-terminale de RAP1 induite par son mode d'interaction avec TRF2 donnerait au complexe de nouvelles spécificités d'interaction avec d'autres partenaires protéiques. Un article publié récemment (Rai et al, 2016) présente des résultats en faveur de la première hypothèse, qui montrent que la région N-terminale de RAP1 est nécessaire pour réprimer la localisation télomérique de SLX4, une protéine interagissant avec TRF2 par un motif TBM (Wan et al, 2013). La seconde hypothèse reste encore à tester.
La région N-terminale de RAP1 contient un motif Y-x-L-x-P décrit dans la littérature comme un motif d'interaction avec le domaine TRFH (domaine de dimérisation) de TRF2 (Chen et al, 2008; Kim et al, 2009) et appelé TBM (TRFH-binding motif). La structure cristallographique du domaine TRFH de TRF2 en complexe avec un peptite de RAP1 contenant le motif Y-x-L-x-P montre qu'il s'agit bien d'un TBM, et la mutation en alanine de ces trois résidus dans RAP1 entière abolit le signal biphasique observé par microcalorimétrie, ce qui confirme l'implication de ce motif dans l'interaction entre les protéines entières. Finalement, les expériences d'empreinte protéique ont montré que l'interaction entre TRF2 et RAP1 engage de vastes surfaces des deux protéines, et que ces surfaces sont plus réduites dans le complexe contenant RAP1 mutée dans le TBM. L'ensemble de nos données est en faveur d'un modèle selon lequel RAP1 lie TRF2 avec une haute affinité par son domaine C-terminal, et peut selon les conditions occuper le site de liaison des motifs TBM dans le domaine TRFH de TRF2 (figure à droite). Ces résultats ont été publiés peu après la fin de ma thèse (Gaullier et al, 2016). Nous avons formulé deux hypothèses (non mutuellement exclusives) sur la fonction de ce mode d'interaction : la première propose qu'un des rôles de RAP1 pourrait être de réguler l'accès au site de liaison des motifs TBM par d'autres protéines contenant ces motifs, la seconde propose que la dimérisation de la région N-terminale de RAP1 induite par son mode d'interaction avec TRF2 donnerait au complexe de nouvelles spécificités d'interaction avec d'autres partenaires protéiques. Un article publié récemment (Rai et al, 2016) présente des résultats en faveur de la première hypothèse, qui montrent que la région N-terminale de RAP1 est nécessaire pour réprimer la localisation télomérique de SLX4, une protéine interagissant avec TRF2 par un motif TBM (Wan et al, 2013). La seconde hypothèse reste encore à tester.Références:
Chen Y, Yang Y, van Overbeek M, Donigian JR, Baciu P, de Lange T & Lei M (2008) A shared docking motif in TRF1 and TRF2 used for differential recruitment of telomeric proteins. Science 319: 1092–1096
Chen Y, Rai R, Zhou Z-R, Kanoh J, Ribeyre C, Yang Y, Zheng H, Damay P, Wang F, Tsujii H, Hiraoka Y, Shore D, Hu H-Y, Chang S & Lei M (2011) A conserved motif within RAP1 has diversified roles in telomere protection and regulation in different organisms. Nat. Struct. Mol. Biol 18: 213–221
Court R, Chapman L, Fairall L & Rhodes D (2005) How the human telomeric proteins TRF1 and TRF2 recognize telomeric DNA: a view from high-resolution crystal structures. EMBO Rep. 6: 39–45
Fairall L, Chapman L, Moss H, de Lange T & Rhodes D (2001) Structure of the TRFH dimerization domain of the human telomeric proteins TRF1 and TRF2. Mol. Cell 8: 351–361
Gaullier G, Miron S, Pisano S, Buisson R, Le Bihan Y-V, Tellier-Lebègue C, Messaoud W, Roblin P, Guimarães BG, Thai R, Giraud-Panis M-J, Gilson E & Le Du M-H (2016) A higher-order entity formed by the flexible assembly of RAP1 with TRF2. Nucl. Acids Res. 44: 1962–1976
Hanaoka S, Nagadoi A, Yoshimura S, Aimoto S, Li B, de Lange T & Nishimura Y (2001) NMR structure of the hRap1 Myb motif reveals a canonical three-helix bundle lacking the positive surface charge typical of Myb DNA-binding domains. J. Mol. Biol. 312: 167–175
Kim H, Lee O-H, Xin H, Chen L-Y, Qin J, Chae HK, Lin S-Y, Safari A, Liu D & Songyang Z (2009) TRF2 functions as a protein hub and regulates telomere maintenance by recognizing specific peptide motifs. Nat. Struct. Mol. Biol. 16: 372–379
Rai R, Chen Y, Lei M & Chang S (2016) TRF2-RAP1 is required to protect telomeres from engaging in homologous recombination-mediated deletions and fusions. Nat Commun 7: 10881
Takai KK, Hooper S, Blackwood S, Gandhi R & de Lange T (2010) In vivo stoichiometry of shelterin components. J. Biol. Chem. 285: 1457–1467 doi:10.1074/jbc.M109.038026
Takai H, Smogorzewska A & de Lange T (2003) DNA Damage Foci at Dysfunctional Telomeres. Current Biology 13: 1549–1556
Sarthy J, Bae NS, Scrafford J & Baumann P (2009) Human RAP1 inhibits non-homologous end joining at telomeres. EMBO J. 28: 3390–3399
Wan B, Yin J, Horvath K, Sarkar J, Chen Y, Wu J, Wan K, Lu J, Gu P, Yu EY, Lue NF, Chang S, Liu Y & Lei M (2013) SLX4 Assembles a Telomere Maintenance Toolkit by Bridging Multiple Endonucleases with Telomeres. Cell Reports 4: 861–869
Le prix Chimie (Morgane Poupon):

Pour obtenir ces nouveaux oxydes de tellure et de sélénium NCs nous avons fait le choix d'utiliser les synthèses par voie hydrothermale basse pression et basse température. Un premier travail de recherche a été réalisé afin de comprendre les mécanismes et les différents paramètres influençant les synthèses hydrothermales (concentration, cycle de température, évolution du nombre d'équivalents de soude, changement de précurseur, etc.). À l'issue de ce travail nous avons pu mettre en place un protocole expérimental, permettant l'étude généralisée de système ternaire M-Te/Se-O (M = Ca, Sr, Ba, Ni, Co) par synthèse hydrothermale. À partir de ces synthèses nous avons suivie deux axes de recherche :
- l'étude structurale de nouveaux polymorphes CaTeO3 obtenus par déshydratations des polymorphes α et β-CaTeO3(H2O) (Figure 1) et par synthèse hydrothermale directe,
- l'étude des structures, des propriétés magnétiques et diélectriques, des composés isotypes Co3Te2O6(OH)2(H2O)0.5 et Ni3Se2O6(OH)2 et des composés hydratés CoSeO3(H2O)x (x = 3/4 et 1/3) et α- et β-NiSeO3(H2O).
La première étude, sur le système Ca-Te-O, nous a permis de mettre en évidence deux nouveaux polymorphes α et β-CaTeO3(H2O), ainsi que quatre nouveaux polymorphes CaTeO3. Les caractérisations des structures ont pu être effectuées à partir de la diffraction des rayons X sur poudre et sur monocristal. Nous avons pu notamment décrire la structure lamellaire désordonnée de la phase β-CaTeO3(H2O) par une structure modulée à l'aide du formalise des superespaces. Parmi ces nouveaux polymorphes trois sont non-centrosymétriques
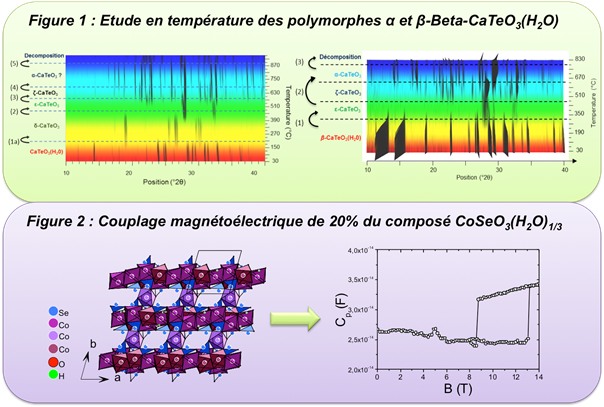 (α-CaTeO3(H2O), δ-CaTeO3 et η- CaTeO3) et l'analyse de leurs propriétés d'optique non linéaire par des mesures de génération de second harmonique, a pu être effectuée. Lors de la deuxième étude, nous avons pu notamment résoudre la structure NC maclée de la nouvelle phase CoSeO3(H2O)3/4 et mettre en évidence la présence d'un couplage magnétoélectrique de l'ordre de 4,5% en dessous de 7K. Néanmoins, c'est pour la phase CoSeO3(H2O)1/3 que nous avons pu mettre en évidence le plus fort couplage magnétoélectrique, qui est de l'ordre de 20% en dessous de 12K. (Figure 2)
(α-CaTeO3(H2O), δ-CaTeO3 et η- CaTeO3) et l'analyse de leurs propriétés d'optique non linéaire par des mesures de génération de second harmonique, a pu être effectuée. Lors de la deuxième étude, nous avons pu notamment résoudre la structure NC maclée de la nouvelle phase CoSeO3(H2O)3/4 et mettre en évidence la présence d'un couplage magnétoélectrique de l'ordre de 4,5% en dessous de 7K. Néanmoins, c'est pour la phase CoSeO3(H2O)1/3 que nous avons pu mettre en évidence le plus fort couplage magnétoélectrique, qui est de l'ordre de 20% en dessous de 12K. (Figure 2)Ce travail de thèse a montré la pertinence de la synthèse hydrothermale pour obtenir de nouvelles phases non-centrosymétriques, dans le cas de l'étude d'oxydes ternaires à base d'éléments à paire d'électrons libres, tels que le tellure et le sélénium (IV). Nous avons également montré que ces phases, lorsqu'elles contenaient des cations magnétiques, étaient de sérieux candidats dans la recherche de matériaux multiferroïques ou présentant un fort couplage magnétoélectrique.
Références:
[1] T. Hahn, International Tables for Crystallography, D. Reidel Publishing Compagny: Dordecht, 1983.
[2] Velev, J. P.; Jaswal, S. S.; Tsymbal, E. Y. Philos. Trans. R. Soc. Lond. Math. Phys. Eng. Sci. 2011, 369 (1948), 3069–3097.
[3] Eerenstein, W.; Mathur, N. D.; Scott, J. F. Nature 2006, 442 (7104), 759–765.
[4] Schmid, H. J. Phys. Condens. Matter 2008, 20 (43), 434201.
[5] Halasyamani, P. S. Chem. Mater. 2004, 16 (19), 3586–3592.
[6] Poupon, Morgane; Barrier, Nicolas; Petit, Sebastien; et al. Inorg. Chem., 2015, 54(12), 5660-5670
Le prix Physique (Maxime Dupraz): Les propriétés physiques à petite échelle de longueur diffèrent fortement de celles du matériau massif, typiquement en deçà du micromètre [1]. Par exemple, la résistance mécanique augmente quand la taille diminue et de fortes contraintes résiduelles liés aux procédés d'élaboration sont présentes au sein de nanostructures [2]. Il existe ainsi un besoin d'une meilleure compréhension de la relation entre la microstructure et les propriétés des matériaux aux échelles sub-micrométriques. La diffraction des rayons-X cohérents (CXD) est une technique émergente de synchrotron qui est très sensible aux champs de déformation et à la présence de défauts structuraux [3]. En principe, une image 3D de la microstructure de l'échantillon peut-être obtenue à partir des données de diffraction cohérente [4]. De plus, les rayons X cohérents peuvent être aussi utilisés pour l'imagerie par holographie de domaines/structures magnétiques [5]. Ces deux techniques sont mises en pratique dans ces travaux de thèse.
Les propriétés physiques à petite échelle de longueur diffèrent fortement de celles du matériau massif, typiquement en deçà du micromètre [1]. Par exemple, la résistance mécanique augmente quand la taille diminue et de fortes contraintes résiduelles liés aux procédés d'élaboration sont présentes au sein de nanostructures [2]. Il existe ainsi un besoin d'une meilleure compréhension de la relation entre la microstructure et les propriétés des matériaux aux échelles sub-micrométriques. La diffraction des rayons-X cohérents (CXD) est une technique émergente de synchrotron qui est très sensible aux champs de déformation et à la présence de défauts structuraux [3]. En principe, une image 3D de la microstructure de l'échantillon peut-être obtenue à partir des données de diffraction cohérente [4]. De plus, les rayons X cohérents peuvent être aussi utilisés pour l'imagerie par holographie de domaines/structures magnétiques [5]. Ces deux techniques sont mises en pratique dans ces travaux de thèse.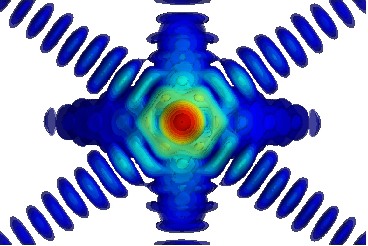 Tout d'abord, nous démontrons que la CXD permet d'identifier quantitativement dans l'espace réciproque tous les types de dislocations, dans le cas d'arrangements simples [6]. Pour des structures plus complexes de défauts, la reconstruction numérique de la densité électronique et de la phase permet de déterminer leur nature et microstructure 3D. Ces deux méthodologies, i.e. l'analyse de la signature des défauts dans l'espace réciproque et la reconstruction dans l'espace réel, sont appliquées au cours d'une expérience de déformation in situ d'une micro-cristallite d'or par nano-indentation. En s'appuyant sur les reconstructions 3D à différents stades de la sollicitation mécanique, la germination une boucle de dislocation prismatique est clairement identifiée. L'interaction entre les défauts germés et la déformation résiduelle dans la cristallite, conduit à une relaxation équivalente à un “recuit mécanique”. De plus la sensibilité de la technique est évaluée dans le cas d'interfaces (surface libre, mâcle, hétéro-épitaxie).
Tout d'abord, nous démontrons que la CXD permet d'identifier quantitativement dans l'espace réciproque tous les types de dislocations, dans le cas d'arrangements simples [6]. Pour des structures plus complexes de défauts, la reconstruction numérique de la densité électronique et de la phase permet de déterminer leur nature et microstructure 3D. Ces deux méthodologies, i.e. l'analyse de la signature des défauts dans l'espace réciproque et la reconstruction dans l'espace réel, sont appliquées au cours d'une expérience de déformation in situ d'une micro-cristallite d'or par nano-indentation. En s'appuyant sur les reconstructions 3D à différents stades de la sollicitation mécanique, la germination une boucle de dislocation prismatique est clairement identifiée. L'interaction entre les défauts germés et la déformation résiduelle dans la cristallite, conduit à une relaxation équivalente à un “recuit mécanique”. De plus la sensibilité de la technique est évaluée dans le cas d'interfaces (surface libre, mâcle, hétéro-épitaxie).
L'holographie magnétique est utilisée pour déterminer les structures magnétiques dans des plots micrométriques de permalloy (FeNi) qui prennent la forme de vortex. De plus leur dynamique sous champ magnétique est résolue en temps (à la nanoseconde).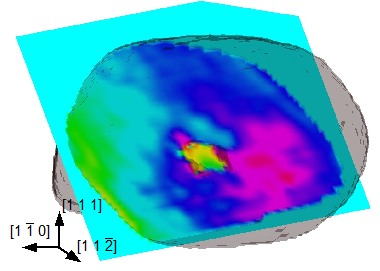
Références:
[1] Uchic et al. (2004), Science 305, 986
[2] Bei et al. (2008), Acta. Mater. 56, 4762-4770.
[3] Robinson & Harder (2009). Nature Materials 8, 291
[4] Newton et al. (2010). Nature Materials 9, 120-124
[5] Eisebitt et ak. (2004) Nature 432, 885
[6] Dupraz et al. (2015). J. Appl. Cryst. 48, 621